ホーム > 砂糖 > 調査報告 > てん菜 > テンサイ西部萎黄病の病原ウイルスと感染源について
テンサイ西部萎黄病の病原ウイルスと感染源について
最終更新日:2018年2月9日
 PDFはこちら
PDFはこちらテンサイ西部萎黄病の病原ウイルスと感染源について
2018年2月
ホクレン農業協同組合連合会 農業総合研究所 営農支援センター
吉田 直人、玉田 哲男
吉田 直人、玉田 哲男
【要約】
近年、北海道各地のてん菜に萎黄性のウイルス病が発生し、大きな被害を与えている。病原ウイルスの解析の結果、これまで病原とされてきたビート西部萎黄ウイルス(米国株)とは生物学的・遺伝学的に異なることが明らかとなり、ウイルス名をビート黄葉ウイルス、病名をテンサイ黄化病へ改名することを提案している。今回は、接種試験から明らかとなった感染源の候補作物や雑草を、本病の伝染環とともに紹介したい。
はじめに
てん菜の萎黄症状の発生経過は明らかでないが、1955年ごろの札幌市周辺の採種地帯における発生が最初の記載とされる。1969年にはビート西部萎黄病ウイルス(beet western yellows virus)が病原ウイルスとして同定され1)、発生生態や防除法について研究が行われた2)、3)。その後、1989〜1992年に西胆振地方と後志地方で激しい黄化症状が発生し4)、一部の地域においては全面に激発した畑もあったことから、急きょ防除対策が求められた5)。その後も各地に散発していたが、2008年以降、てん菜の主要生産地である十勝地方・網走地方で大発生、甚大な被害を与えたことから再び注目され、多発原因の究明と対策が要望されている。
本病の病原であるビート西部萎黄ウイルスは世界中に多くの系統が分布し、さまざまな作物に感染する6)。北海道のてん菜に発生するウイルスは米国の西部萎黄ウイルスに近縁であるとの報告7)はあるが、ウイルス系統の存在や寄生性など、不明な点が多い。そこでわれわれは、病原ウイルスについて、遺伝子解析と併せて特に宿主範囲について詳細に実験を行った。
本病の病原であるビート西部萎黄ウイルスは世界中に多くの系統が分布し、さまざまな作物に感染する6)。北海道のてん菜に発生するウイルスは米国の西部萎黄ウイルスに近縁であるとの報告7)はあるが、ウイルス系統の存在や寄生性など、不明な点が多い。そこでわれわれは、病原ウイルスについて、遺伝子解析と併せて特に宿主範囲について詳細に実験を行った。
1.病徴と発生状況について
ウイルス感染による黄化症状は7月中下旬から見られることがあるが、通常は9〜10月に症状がより明瞭となる。下葉(古い葉)から中葉にかけ特に葉脈と葉脈の間が黄化し、古い葉は厚みを帯びてゴワゴワした感じになる(写真1)。てん菜の畑では、黄化症状が数株から数十株単位でスポット状に発生する場合が多い(写真2)が、2014年の十勝地方では、写真3のように全面が黄化した畑も多数見られた。このような発病をすると糖量で30%程度も減収する5)。



2.病原ウイルスについて
当初、本病はビート西部萎黄ウイルスによって起こり、病名はウイルス名から西部萎黄病とされた1)。米国西部で発見されたことにちなみ、病名にも“西部”が入る。宿主範囲が極めて広いことで知られ、23科150種以上の植物への感染の報告がある6)。また、寄生性や病徴が異なる複数の系統が存在するとされるが、北海道における詳細な調査は行われていなかった。そこで、われわれはウイルスの寄生性および遺伝的な側面から、北海道で発生するウイルスの特徴付けを行った。その結果、北海道で西部萎黄病とされてきた病害には、少なくとも2種のウイルスが関与し、米国の西部萎黄ウイルスとはいずれも異なることが明らかとなった。これを踏まえ、てん菜に萎黄症状を引き起こすものを“ビート黄葉ウイルス(beet leaf yellowing virus)”、また病名を“テンサイ黄化病”へと変更することを提案し8)、病名委員会で承認された。このウイルスの大きな特徴としては、西部萎黄ウイルスの宿主であるアブラナ科作物(はくさい、かぶ、キャベツ、ブロッコリー、カリフラワー、だいこんなど)には感染しない点である9)、10)。もう一方のウイルスは、ほうれんそうなどを共通の宿主とするものの、てん菜には感染しないことが分かり、アブラナ科作物に広く存在する“アブラナ萎黄ウイルス(brassica yellows virus)”と同定した9)、10)。なお、西部萎黄ウイルス検出用として市販されている2種のELISAキット(LOEWE社、Agdia社)によりこれらの2種ウイルスを容易に検出できるが、識別が困難であるため、混乱しないよう注意が必要である11)。
3.アブラムシによるウイルスの伝播ついて
ウイルスは直径25ナノメートルの球状で、モモアカアブラムシによって伝播される。アブラムシは罹病植物を1時間以上吸汁するとウイルスを保毒し、約1日の潜伏期間を経て、最短10分の吸汁でウイルスをうつす。なお、吸汁時間が長いほど伝染率は高まる。一度保毒した個体は、一生ウイルスを感染させる能力を持つが、子孫(子虫)には伝わらない。
モモアカアブラムシは、体長1〜2ミリメートル、卵円形で、体の色は黄緑色、緑色、赤褐色など変化がある(写真4)。触覚の長さは体長とほぼ同じで、尾片は体と同色となり、角状管の先端は暗色である。アブラムシは、越冬寄主のバラ科植物上で秋期に産卵し、卵の状態で越年した後、春にふ化して幹母となる。北海道では、地域によって異なるが、6月上旬ごろから有翅虫が出現し、てん菜圃場での発生は7月から8月にピークを迎える(図)。しかし、ハウス内で成虫が越冬した場合には、発生時期が早まることが予想され、春期ハウス内や近傍の雑草や野菜類で繁殖し、そこから出現した有翅アブラムシがてん菜畑に飛来する。
モモアカアブラムシは、体長1〜2ミリメートル、卵円形で、体の色は黄緑色、緑色、赤褐色など変化がある(写真4)。触覚の長さは体長とほぼ同じで、尾片は体と同色となり、角状管の先端は暗色である。アブラムシは、越冬寄主のバラ科植物上で秋期に産卵し、卵の状態で越年した後、春にふ化して幹母となる。北海道では、地域によって異なるが、6月上旬ごろから有翅虫が出現し、てん菜圃場での発生は7月から8月にピークを迎える(図)。しかし、ハウス内で成虫が越冬した場合には、発生時期が早まることが予想され、春期ハウス内や近傍の雑草や野菜類で繁殖し、そこから出現した有翅アブラムシがてん菜畑に飛来する。

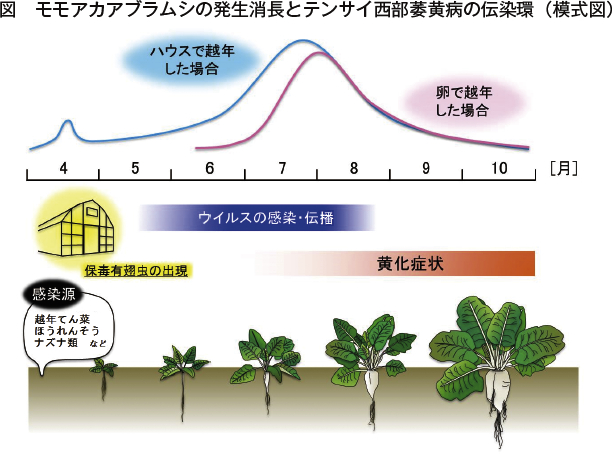
写真2のような大型の発病は、早期に保毒した有翅虫が、一株(または移動して数株)に感染を起こし、そこに生じた無翅虫によって感染が拡大したものである。圃場中のスポットの数が飛来した個体数を反映しており、スポットのサイズは、そこで増殖した個体による二次的な感染程度を示している。
4.感染源について
本病の発生生態への理解を深める上で、病原ウイルスの宿主範囲を知ることは極めて重要となる。そこでわれわれは、17科100種以上の植物を用いた接種試験を行い、7科26種の植物に感染することを明らかにした。そのうち感染源となり得る候補植物種を表に示す。前述の通り、これまで宿主とされてきた多くのアブラナ科の作物は実際には感染せず、イヌガラシ属(スカシタゴボウ、ヤチイヌガラシ)、タネツケバナ属(タネツケバナやミチタネツケバナ)、ハルザキヤマガラシなどの道内に広く分布し、畑地・空き地・道端に生育する雑草も、ウイルスに感染しないことが分かった。一方、越年した黄化症状を呈するナズナからは実際にウイルスが検出された(写真5)。


また、過去には越冬ハウス内でほうれんそうやアブラナ科作物が残存し、生育しているのが観察された(写真6)。アブラナ科の作物や雑草はモモアカアブラムシの嗜好性が高く、開花期には花器部やその茎部に群生することから、本病の発生源になり得る可能性が高い。

おわりに
本病に対する有効な対策を講じるためには、発生生態、特に感染源とアブラムシの発生時期(消長)を知る必要がある。これまで、北海道の西部萎黄ウイルスは米国株と同様に宿主範囲が広いとされてきたが、本試験の結果、宿主範囲はむしろ狭く、限られた植物にしか感染しないことが分かった。実際にどの植物が自然界でどの程度感染し、感染源としての役割を担っているかについては、さらに検討していきたい。
北海道におけるモモアカアブラムシの発生時期は通常では6月ごろとされているが、過去の調査5)や最近の調査12)により、越年ハウス内でアブラムシや感染植物が生存することが明らかとなった。このことから、アブラムシが通常より早い時期から発生し、さらに感染源からウイルスを獲得できるような条件が整ったことにより、近年の大発生が起こったものと考えられる。本病を効果的に防除するためにはアブラムシが越年できないよう、越冬ハウス内の植物を除去するなど、適正な管理が重要となるだろう。
謝辞
本研究で用いたヒメアマナズナ、ツノミナズナ、クジラグサの3種の種子は、長野県農業試験場の青木政晴氏により分譲されたものである。
北海道におけるモモアカアブラムシの発生時期は通常では6月ごろとされているが、過去の調査5)や最近の調査12)により、越年ハウス内でアブラムシや感染植物が生存することが明らかとなった。このことから、アブラムシが通常より早い時期から発生し、さらに感染源からウイルスを獲得できるような条件が整ったことにより、近年の大発生が起こったものと考えられる。本病を効果的に防除するためにはアブラムシが越年できないよう、越冬ハウス内の植物を除去するなど、適正な管理が重要となるだろう。
謝辞
本研究で用いたヒメアマナズナ、ツノミナズナ、クジラグサの3種の種子は、長野県農業試験場の青木政晴氏により分譲されたものである。
参考文献
1)讃井蕃、村山大記(1969)「サトウダイコン西部萎黄病ウイルス(Sugar beet western yellows virus)について」『日植病報』(35)p.125(講要).
2)杉本利哉、村山大記、讃井蕃(1970)「てん菜の萎黄性ウイルスに関する研究(第1報)」『てん菜研究報告』(8)pp.1-163.
3)杉本利哉、村山大記(1972)「てん菜の萎黄性ウイルス病に関する研究(第2報)」『てん菜研究報告』(13)pp.1-72.
4)成田正孝、菅原寿一、木村雅暢、楠目俊三、玉田哲男(1992)「西胆振・後志地方における西部萎黄病の多発について」『てん菜研究報』(34) pp.133-1138.
5)北海道中央農業試験場(1994)「テンサイ西部萎黄病の多発原因の究明と対策」『平成5年度農業試験会議(成績会議)資料』pp.1-38.
6)Duffus, J. E.・Johnstone, G. R.(1983)「Beet western yellows luteovirus」『Plant Viruses Online』(http://sdb.im.ac.cn/vide/descr089.htm)
7)上田重文、三宅規文、安岡真二、池谷美奈子、高橋宙之(2014)「北海道で発生するビート西部萎黄ウイルスの系統解析」『北日本病虫研報』(65)pp.37-41.
8)吉田直人、丹羽昌信、玉田哲男(2017)「北海道産ビート西部萎黄ウイルス(BWYV)分離株の特徴:ウイルス名の変更について」『日植病報』(83)p.74(講要).
9)吉田直人、丹羽昌信、玉田哲男(2016)「北海道のテンサイおよびアブラナ科作物に感染する複数のポレロウイルスについて」『日植病報』(82) pp.73-74(講要).
10)吉田直人、玉田哲男(2017)「北海道に発生するビート黄化ウイルス(BLYV)とアブラナ萎黄ウイルス(BrYV)の寄主範囲と遺伝的特性」『日植病報』(83)p.215(講要).
11)吉田直人、丹羽昌信、玉田哲男(2016)「テンサイ西部萎黄病に関与するウイルスの寄生性、エライザ検出および分類学的考察」『てん菜研究会』 (57)pp.31-33.
12)三宅規文(2017)「テンサイの西部萎黄病の効果的抑制方法と十勝全域における大規模検証」『平成29年度日本植物病理学会北海道部会年報』(45)pp.32−33.
1)讃井蕃、村山大記(1969)「サトウダイコン西部萎黄病ウイルス(Sugar beet western yellows virus)について」『日植病報』(35)p.125(講要).
2)杉本利哉、村山大記、讃井蕃(1970)「てん菜の萎黄性ウイルスに関する研究(第1報)」『てん菜研究報告』(8)pp.1-163.
3)杉本利哉、村山大記(1972)「てん菜の萎黄性ウイルス病に関する研究(第2報)」『てん菜研究報告』(13)pp.1-72.
4)成田正孝、菅原寿一、木村雅暢、楠目俊三、玉田哲男(1992)「西胆振・後志地方における西部萎黄病の多発について」『てん菜研究報』(34) pp.133-1138.
5)北海道中央農業試験場(1994)「テンサイ西部萎黄病の多発原因の究明と対策」『平成5年度農業試験会議(成績会議)資料』pp.1-38.
6)Duffus, J. E.・Johnstone, G. R.(1983)「Beet western yellows luteovirus」『Plant Viruses Online』(http://sdb.im.ac.cn/vide/descr089.htm)
7)上田重文、三宅規文、安岡真二、池谷美奈子、高橋宙之(2014)「北海道で発生するビート西部萎黄ウイルスの系統解析」『北日本病虫研報』(65)pp.37-41.
8)吉田直人、丹羽昌信、玉田哲男(2017)「北海道産ビート西部萎黄ウイルス(BWYV)分離株の特徴:ウイルス名の変更について」『日植病報』(83)p.74(講要).
9)吉田直人、丹羽昌信、玉田哲男(2016)「北海道のテンサイおよびアブラナ科作物に感染する複数のポレロウイルスについて」『日植病報』(82) pp.73-74(講要).
10)吉田直人、玉田哲男(2017)「北海道に発生するビート黄化ウイルス(BLYV)とアブラナ萎黄ウイルス(BrYV)の寄主範囲と遺伝的特性」『日植病報』(83)p.215(講要).
11)吉田直人、丹羽昌信、玉田哲男(2016)「テンサイ西部萎黄病に関与するウイルスの寄生性、エライザ検出および分類学的考察」『てん菜研究会』 (57)pp.31-33.
12)三宅規文(2017)「テンサイの西部萎黄病の効果的抑制方法と十勝全域における大規模検証」『平成29年度日本植物病理学会北海道部会年報』(45)pp.32−33.
このページに掲載されている情報の発信元
農畜産業振興機構 調査情報部 (担当:企画情報グループ)
Tel:03-3583-8678
農畜産業振興機構 調査情報部 (担当:企画情報グループ)
Tel:03-3583-8678